Background
In Sudan, malaria is a major health problem heading the list of the diseases afflicting the country. Of the seven recognized species of the Anopheles gambiae complex, An. arabiensis Patton is the most abundant and most important vector of human malaria in Sudan. It has been regarded from many localities in Sudan, from the extreme south up to the northern borders with Egypt. It was reported from many localities in Kassala State, eastern Sudan by (Lewis, 1959; Haridi, 1972; Petrarca et al., 1986; Himeidan et al., 2004; Hamza et al., 2005; 2014).
The morphological description of this mosquito has been given by
Evans (1938), cited in
Gillies and De-Meillon (1968) and
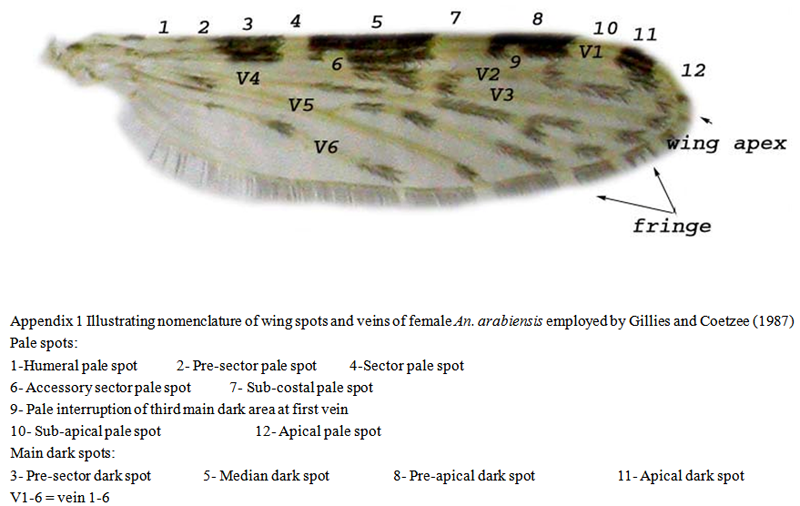
Gillies and Coetzee (1987).Morphological characters appear to be in many cases a very useful tool for anopheline taxonomy. Morphometrics has proved useful in studying some species complexes, when used in conjunction with statistical analysis (
Coluzzi, 1964;
White, 1977;
Lambert and Coetzee, 1982;
Service, 1988). Morphometrics concerned with variations and changes in the body form of an organism. It transforms the complex body forms into quantitative series of numbers that can be analyzed and used for comparisons of different forms (
Daly, 1985).
Since the discovery that
An. gambiae is not a single species but a complex of cryptic species, many workers attempted to find morphological differences between the members of the complex including
An. arabiensis (
Coluzzi, 1964;
Ramsdale and Leport, 1967). Subsequently, many workers have attempted to find morphological variations between the three fresh water species-
An. arabiensis,
An. gambiae and
An. quadriannulatus (
Ismail and Hammoud, 1968;
Zahar et al., 1970;
Clarke, 1971;
White and Muniss, 1972) without success.
Petrarcaet et al. (1998) carried out a morphometric analysis on
An. arabiensis and
An.
gambiae from different sites of Sub-Saharan Africa and he found that all the measures were significantly larger for
An. arabiensis.
Ribeiroet et al. (1979) described
An. quadriannulatus davidsoni, a member of the
An.
gambiae group based on morphological criteria only.
Schmidtet et al
. (2003) proposed taxonomic identification keys for the members of the
An. punctulatus complex based on morphological features of the adults and immature stages. The separation between the species of the
An.
gambiae complex is much less reliable owing to the existence of a considerable overlap between the different character distributions (
Bryan, 1980;
Bushrod, 1981;
Coetzee, 1989).
The present study was conducted to characterize the morphology of adult females An. arabiensis found in eastern Sudan and to investigate the existence of morphometric variations among local populations of this important malaria vector. Relatively few studies have addressed this important vector species in eastern Sudan. However, field based studies such as that have much to add to our understanding of malaria transmission in the area. The results of the study can help in the correct identification of this malaria vector. Correct identification of malaria vectors is essential for the correct evaluation of malaria vector ecology studies and control programs in the area. The results of the morphological measurements of An. arabiensis revealed in this study can be used with the published morphological data of the members of An. gambiae for proposing morphological identification key for the complex.
1 Materials and Methods
1.1 Collection sites and mosquitoes used for the morphometric study
Two hundred and fourteen field samples of adult females
An. arabiensis mosquitoes were collected from four sites in Kassala State (Kassala (52 individuals), New Halfa (52 individuals), Aroma (54 individuals) and Wager (56 individuals)). The state lies in the eastern part of the Sudan between 34°: 40˝ & 37° E and 14°: 45˝ & 17°: 40˝ N, (
Figure 1) under arid and semi-arid climate with rainfall of varying intensity and duration. The climate in the state is characterized by three seasons, the short rainy season (July-October), cool dry season (November-February) and hot dry season (March-June).
The topography of the state is characterized by high-lands to the East and South and flat plains to the West and North. Atbara and Gash rivers represent the most important water courses in the state. The Atbara River is flowing out of Ethiopia and it is the largest seasonal river in the state, and the only one which has a discharge during the greatest part of the year (July- December). The Gash River originates in Eritrean high-lands and flows across the flat plain from late June to October. About 10 Km North of Kassala town, the Gash River fans out in an inland delta, the Gash Delta.
Kassala town (15°: 28˝ N & 36°: 24˝ E) is located in an area characterized by the presence of the back-bones of mountains surrounding the eastern part in addition to presence of the horticultural farms of vegetables and fruits along the Gash river banks. New Halfa town (15°: 19 ˝N & 35°: 36˝ E) is located in the western bank of the Atbara River in the middle of New Halfa an agricultural scheme. Aroma (15°: 55˝ N & 36°: 11˝E) and Wagar (16°: 09˝ N & 36°: 12˝E) localities are located in flat area with a moderate vegetation cover in the northern part of Gash Delta. The major consideration in the selection of the study sites is that they represent the different environmental conditions of the state. The selection is also based on the easy accessibility of the collection sites.
For comparison, specimens of females An. arabiensis (52 individuals) were obtained from laboratory colonies- originating from Sennar State (34°: 3˝ and 34°: 05˝ E & 12°: 58˝ and 12°: 97˝ N, central Sudan) and maintained at Sennar town (13°: 33˝ N & 33°: 37˝ E) since 2007- were also embedded in the study.
1.2 Samples collection and preservation
In door resting wild females
Anopheles mosquitoes were caught from rooms by hand capture (
WHO, 1975) during the rainy season 2008. Collected mosquitoes were fixed alive in the field in 70% ethanol and preserved individually at -20 °C for subsequent processing in the laboratory. The processing of the materials for this study was performed at the Department of Zoology, Faculty of Science, and University of Khartoum, Sudan.
1.3 Identification and mounting offemales An. arabiensis
Members of the
An. gambiae complex were sorted out of other anopheline mosquitoes using morphological identification keys described by
Gillies and De-Mellion (1968) and
Gillies and Coetzee (1987) with the aid of the dissecting microscope. Subsequently, specimens were dissected out, the head, wings and legs were carefully detached from the body and mounted individually in Puri's mounting medium on glass microscopic slides as described by
WHO (1975) with minor modifications. Mosquito body parts were mounted directly into Puri's mounting medium without prior clearing because gum-chloral mountant continues to clear samples after mounting. The remaining of the dissected parts were preserved in 70% alcohol and stored at -20 °C for molecular identification. Females
An. arabiensis were identified by molecular polymerase chain reaction (PCR) techniques using the protocol developed by
Scott et al. (1993). For morphometric analysis, the permanent slides were used for detailed examination of the external body structures of
An. arabiensis.
1.4 Morphological characters measurements
First, a preliminary list was prepared from all morphological characters of the adult female An. arabiensis. Out of this list, all characters that fixation could not have changed were selected. Thirty seven metric characters were measured on 270 females An. arabiensis represented all the collection sites.The characters consisted of one character associated with antennae, 10 characters with palps, 14 characters with wings and 12 characters of the legs (Table 1).
Morphological measurements were done by the projection method adopted by (
Zahar et al., 1970). Selected characters were measured using a Wild MII binocular calibrated compound microscope fitted with and 1.25X phototube. The slide mounted specimens were projected with a phototube on a microscope field using 3.5X objective and 8X eyepiece. After an excellent view of the projected image was obtained, the projected image was drawn and subsequently measured to the nearest half millimeter. Then the measures obtained were calibrated to the real lengths using a micrometer stage (1mm-Erma, Tokxc).
1.5 Data analysis
All statistical analysis of the morphometric data were conducted using SPSS Version 16.0 for Windows. The morphological measurements were not transformed to ratios so as to preserve the possible influence of differences in the body sizes of the species.
The measured data were subjected to univariate and multivariate statistical analysis.Univariate statistics involved calculation of descriptions, One- Way ANOVA testand Box – and whisker plots. Descriptions involvedcalculation of mean size measurements plus or minus their standard error and sample sizes. A One- Way ANOVA testwas carried out to test the significant differences of the measured characters among different populations of An. arabiensis. Box – and whisker plots were used to check for the presence of outliers and to summarize the univariate data.
Multivariate statistics such as, principal component analysis, discriminant function analysis and cluster analysis were used.
Principal component analysisis a method of data reduction and it was used to simplify subsequent analysis of the morphological data of field specimens. The analysis aims to produce a small number of derived variables that explain most of the variance and can be used in place of the large numbers of original variables. It obtains a set of principal components by weighting all the available variables. The first component explains the most variation; the second explains the next most variation, and so on. Investigation of the first few components will show which variables contribute most to the variations between individuals (
Dythan, 2003;
Pimentel, 1992).
Discriminant function analysis concerns with classification and used to obtain a small number of useful discriminating variables. Since the first component of the principal component analysis gave rise to 68.4% of the original variance, it was used as new variables to carryout multivariatediscriminant function analysis to discriminate between the different field populations of An. arabiensis.
Cluster analysis concerns with classification and it is essentially a data reduction tool that creates subgroups that are more manageable than individual’s datum. The output of the analysis is shown as a dendrogram of relationships among groups. Cluster analysis (unweighted pair-group method, arithmetic averages) was used to examine the relationship of morphological similarity between the different populations of An. arabiensis.
2 Results
2.1 Morphological measurements
The mean, standard error and sample size of the measured morphological characters on five populations of females
An. arabiensis were listed in
Table 1.
2.2 Investigation of morphometric variations
2.2.1 One- Way ANOVA test
The results of this test as presented in
Table 1 showed that 92% of all the measured characters showed highly statistically significant differences (P<0.05) among all the populations studied. The populations were not significantly different in the following characters: humeral pale spot, sector pale spot and pale interruption of the third main dark area of 1
st vein of the wing. There was a notable overlap in most of the characters ranges.
2.2.2 Principal component analysis
Table 2 shows the results of principal component analysis which was conducted on the measurements of the four field populations. The test simplified the analysis of
Table 1 and showed that three components with Eigen values more than 1 explained the data. Component one contributed the highest Eigen value of 0.
43 with a percentage variance of 68.
4%. It was largely influenced by the length measures of legs and wings (measurements of the mosquito body size). Component 2 and 3 showed Eigen values and percentages variance of 0.
09,
14.
2% and 0.
06, 9.
6%, respectively. The variables of these two components were less important due to the lower percentages of variance contributed by their components. Together, component 1, 2 and 3 account for 9
2.
2% of the total variance.
Box –and whisker plots (
Figures 2&
3) summarize the scores of component 1
and component 2. The scores of components 1 and 2 showed statistically significant differences (P<0.05) between all the four populations studied. The inspection of principal component 1 showed that An. arabiensis populations from Kassala and Aroma town was significantly different from other populations and had the largest body size, followed by the population from New Halfa. Wagar individuals had the smallest mean body size, as shown by the smaller average principal component.
2.2.3 Discriminant function analysis
The analysis was applied on the characters linked with the mosquito body size to discriminate between the four field populations of
An. Arabiensis (
Figure 4). These characters comprised the eight most influential characters for component 1, derived by the principal component analysis (
Table 2). The characters used were: mid leg length, hind leg length, wing length excluding the fringe, wing width excluding the fringe, fore leg length, length of palpal segment III, antenna flagellum length and length of palpal segment IV.
When these characters were subjected to discriminant function analysis, three significant functions were derived.
Table 3 shows the standardized, unstandardized and loading coefficients for the discriminant functions calculated from the 8 selected variables. The first function which accounts for 53.4% of the variance (Wilks Lambda=0. 077; Chi-square (
χ2) =56.28; df= 24; P=0.000) with wing size measurements having high discriminant loadings. This function discriminate Kassala populations from the remaining populations. The second function which accounts for 24.6% of the variance (Wilks Lambda=0.253; Chi-square (
χ2) =30.24; df=14; P=0.007) discriminate New Halfa and Aroma populations from Kassala and Wagar. The third function which accounts for 22% of the variance (Wilks Lambda=0.517; Chi-square (
χ2) =14.50; df=6; P=0.025) discriminate New Halfa population from the other populations (
Table 4). However, this discrimination was not a complete one, since only 37.9% of the original grouped specimens, and 64.5% of the cross-validated ones were correctly classified (
Table 5).
2.2.4 Cluster analysis
Cluster analysis based on a matrix of distance for 37 morphological characters (
Table 1) and complete linkage between groups was used to produce hierarchical clusters of the field and colony populations in a distance dendogram (
Figure 5).
The cluster shows that field samples of An. arabiensis clustered together, with 2 subclusters, one containing population Kassala and Aroma and the other containingpopulation New Halfa and Wagar. The laboratory colony specimens clustered alone and well apart because of their smaller body sizes. The dendogram confirms the results obtained by principal component analysis and showed that the clustering of all the populations based mainly on the measurements of the mosquito body size.
3 Discussion
Measurements of various morphological characters of females
An. arabiensis from Sudan were in accordance with the published data of the same species from other sites along the distribution range of the species (
Coluzzi, 1964;
White et al
.; 1972;
Petrarca et al
., 1998;
Adeiekeet al
., 2008).
Principal component analysis showed that the populations studied differed mainly in the mosquito body size. Kassala and Aroma populations had bigger body size followed by New Halfa. Wagar population had the smallest body sizes. The results confirmed the report of (
Dythan, 2003) on the output of principal component analysis. In this morphometric analysis, it is usually the case that individual specimens will vary in size. The first principal component will always account for size and it is often employed as a method for removing size from the analysis. The second and the rest of the principal components are usually interpreted as shape component (
Dythan, 2003). Unlike the first component, which is positively contributed to all dimensions, the second principal component as well as other components are positively correlated to some variables, and negatively correlated to others.
Based on the results of the morphometrics analysis, the geographical features characterizing the different collection sites may influence the microclimatic conditions of the area which had a direct effect on the mosquito body sizes. The results drawn from this study indicate the presence of some geographical variations among local
An. arabiensis populations. This suggests the presence of some degree of biogeographical structuring within the species range which may be correlated with the environmental conditions persisting in these localities.
Wasserberget al. (2003) mentioned that difference in morphometrics measurements may be attributed to the effect of microclimatic conditions within the microhabitat niche of the species that affect the species abundance, adult life span and larval development.
Thompson (1970) and
Petrarca et al. (1998) reported obvious variations on the mosquito body sizes among field collected females
An. arabiensis and
An.gambiae. They stated that the larger body size of females
An. arabiensis could be caused by the cooler climate of the collection sites in comparison to the other warm collection sites. Moreover, the influence of temperature on the body size (expressed as an inverse, temperature- size correlation) has been repeatedly reported in mosquitoes as
An. merus (
Ie Sueuret al
., 1992) and
An. crucians (
Huet al
., 1993).
Results of various biological data on adult mosquitoes over the years had revealed differences in characteristics of some species depending on the ecotypes and the season (
Lehane, 1991). The host seeking behavior and vectorial capacity of some members of
An. gambiae differ significantly in different geographical zones of the world (
Adeleke, 2008).
A discriminant function analysis was carried out to select morphological characters which could differentiate local populations of females An. arabiensis. Using the body size measurements, the analysis revealed that geographical clustering of field females An. arabiensis populations was not clear and the body size characters had little discrimination values.
Cluster analysis was carried out using the complete set of measured characters which were subjected to principal component analysis. The analysis summarized the relationships among all the populations of An. arabiensis according to their mean body sizes. As shown in the cluster analysis results, An. arabiensis individuals obtained from Sennar laboratory colony became an out group and well apart because of their smaller body sizes. Kassala and Aroma populations appeared as sisters group because of their larger body sizes, while New Halfa and Wagar populations clustered together.
An. arabiensis females obtained from colony materials had a smaller mean body size compared to the field collected ones. This noticeable decrease in the body size of the laboratory specimens may be a result of stressful environmental influences, such as larval crowding or high mean temperature or both, which can lead to a decrease of the body size (
Bock and Milby, 1981). Moreover,
Coluzzi (1964) investigated the morphological characters of members of
An. gambiae complex obtained from colony materials. He stated that, there may have been changes in the mosquito phenotypes as a result of forced gene exchange or because of the selection pressure by artificial conditions.
Munstermann (1994) and
Petrarca et al. (1998) mentioned that the colonization process can modify the organism phenotype, which can make conclusions drawn from observations on laboratory populations’ unreliable.
The results of the study can aid in the description of detailed morphology of adult female An. arabiensis. The morphometric analysis confirmed the existence of some geographical variations in the mosquito body size among local populations of An. arabiensis in eastern Sudan. These variations may be effected with the environmental conditions characterized the localities.
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
Hamza A. M. carried out the technical work and drafted the manuscript. Abukashawa S. M. A contributed to the reading and revising the manuscript. El Rayah El-A. M. supervised the research group and participated in revising the draft manuscript. All authors read and approved the submitted version of the manuscript.
Acknowledgements
The authors thank Dr. Noteila Mustafa Khalid, Department of Zoology, Faculty of Science, Khartoum College of Medical Sciences for her collaboration and assistance in the statistical analysis of the results. We thank Mr. Abd El monem Faza, Director of Dr. El Gadal Center, Sennar, Federal Ministry of Health for providing the laboratory colony materials of An. arabiensis. We thank Department of Zoology, Faculty of Science, and University of Khartoum for the institutional support and use of the facilities of the Zoology Laboratory. This study was supported by a grant from Kassala University.
Adeieke M.A., Mafiana C.F., Idowu A.B., Amusan A.A., and Adekunle M.F., 2008, Morphometric studies on Anopheles gambiae complex (Diptera: Culicidae) in Abeokuta, southwest Nigeria, Acta Entomolegica Sinica, 51(12): 1289-1292
Bock M.E., and Milby M.M., 1981, Seasonal variation of wing length and egg raft size in Culex tarsalis, Proc Calif Mosq, Vector Control Assoc, 49: 64-66
Bryan, J.H., 1980, Use of palpal ratio and the number of pale bands on the palps in separating Anopheles gambiae Giles s. s. and Anopheles melas Theobald (Diptera: Culicidae), Mosq. Syst., 12: 155-163
Bushrod F.M., 1981, The Anopheles gambiae Gilescomplex and Bancroftian Filariasis transmission in a Tanzanian coastal village, Ann. Trop. Med. Parasitol., 75: 93-100
Clarke J.L., 1971, Potential use of spermatheca in the separation of species A and B females of the Anopheles gambiae complex in northern Nigeria, Bull WHO, 45: 260-263
Coetzee M., 1989, Comparative morphology and multivariate analysis for the discrimination of four members of the Anopheles gambiae group in southern Africa, Mosquito Systematics, 21(2): 100-116
Coluzzi M., 1964, Morphological divergence in the Anopheles gambiae complex, Riv. Malariol., 43: 197-232
Daly H.V., 1985, Insect morphometrics, Annual Review of Entomology, 30: 415-438
Dythan C., 2003, Choosing and using statistics: A Biologist/s Guide 2, Malden, Massachusetts, USA: Blackwell Publishing
Evans A.M., 1938, Mosquitoes of the Ethiopian Region, British Museum, National History
Gillies M.T., and Coetzee M., 1987, A supplement to the Anophelinae of Africa South of the Sahara (Afrotropical region), South African Institute for Medical Research, Johannesburg, South Africa
Gillies M.T., and De-Mellion B., 1968, The Anophelinae of Africa South of the Sahara, South African Institute for Medical Research, Johannesburg, South Africa
Hamza A.M., El-Rayah El-A., and Abukashawa S.M., 2014, Molecular characterization of mosquitoes of Anopheles gambiae species complex (Diptera: Culicidae) from Sudan and Republic of Southern Sudan, Journal of Mosquito Research, 4 (13): 1-10
Hamza A.M., Himeidan Y.El-S., Adam I., El-Rayah El-A., 2005, The ecology of Anopheles arabiensis and insecticide resistance/ susceptibility status in Kassala area, eastern sudan, Gezira Journal of Health Sciences, 2(2)
Haridi A.M., 1972, Partial exophily of Anopheles gambiae species B in Khashm El-Girba area in eastern Sudan, Bull Wld Hlth Org, 46: 39-46
Himeidan Y.El-S., Dukeen M.Y., El-Rayah E-lA., and Adam I., 2004, Anopheles arabiensis: abundance and insecticide resistance in an irrigated area of eastern Sudan, Eastern Mediterranean Health Journal,10: 167-174
Hu G.Y., Lounibos L.P., Escher R.L., 1993, Seasonal abundance, age composition and body size of salt-marsh Anopheles (Diptera: Culicidae) in South Florida, J Med Entomol, 30: 883-887
Ie Sueur Sharp D.B. and Appleton C.C., 1992, Dark-scaled areas on adult Anopheles mosquitoes are selectively affected by temperature-related size variation, Med. Vet. Entomol., 6: 396-398
Lambert D.M. and Coetzee M., 1982, A dual genetic and taxonomic approach to the resolution of the mosquito taxon, Anopheles (Cellia) marshalli (Culicidae), Systematic Entomology, 7: 320-332
Lehane M.J., 1991, Biology of blood sucking insects, Herperollins Academic London, pp.288
Lewis D.J., 1959, The anopheline mosquitoes of Sudan, Bull Ent Res, 47: 475-494
Munstermann L.E., 1994, Unexpected genetic consequences of colonization and inbreeding: allozyme tracking in Culicidae (Diptera), Ann Entomol Soc Am, 87: 157-164
Petrarca V., Nugud A.D., Ahmed M.AH., Haridi A.M., Abd-El-Nur O.M., and Coluzzi M., 1986, Dati Preliminari sul complesso Anopheles gambiae in Sudan, Parasitologia, 28: 304-306
Petrarca V., Sabatinelli G., Toure Y.T., and Di Deco M.A., 1998, Morphometric multivariate analysis of field samples of adult Anopheles arabiensis and Anopheles gambiae s. s (Diptera: Culicidae), J. Med. Entomol., 35 (1): 16-25
Pimentel R.A., 1992, An introduction to ordination, principal components analysis and discriminant analysis, In JT Sorensen, R Foottit (eds), Ordination in the Study of Morphology, Evolution and Systematics of Insects, Elsevier Science Publisher, Amsterdam, 11-28
Ramsdale, C.D., and Leport G.H., 1967, Studies of the Anopheles gambiae complex in West Africa, Bull WHO, 36: 494-500
Ribeiro H.C., Ramos C.A., Capela R.A, 1979, Description and biometric study of Anopheles (Cellia) quadriannulatus davidsoni ssp, a seventh member of the Anopheles gambiae Giles complex (Diptera: Culicidae) endemic to the Cape Verde Archipelago, Garcia de Orta Ser Zool, 8: 75-88
Schmidt E.R., Foley D.H., Burgoro H., Bryan J.H., 2003, A morphological study of the Anopheles punctulatus group (Diptera: Culicidae) in the Salomon Islands, with adscription of Anopheles (Cellio) irenicus Schmidt, spn, Bulletin of Entomological Research
Scott J.A., Borgdon W.C., and Collins F.H., 1993, Identification of single specimen of Anopheles gambiae complex by the polymerase chain reaction, American Journal for Tropical Medicine and Hygiene, 49: 520-529
Service M.W., 1988, New tools for old taxonomic problems in blood sucking insects in Biosystematics of haematophagous insects, ed. (M. W. Service), 325, Clarendon press box
Thompson B.W., 1970, The climate of Africa. Oxford University Press, London
Wasserberg G., Yarom I., andWarburgy A., 2003, Seasonal abundance patterns of the sand fly Phelebotomas papatsi in climatically distinct foci of cutaneous Leishmaniasis in Israeli deserts, Medical and Veterinary Entomology, 17: 452-456
White G.B., 1977, The place of morphological studies in the investigation of Anopheles species complexes, Mosq Syst, 9: 1-24
White G.B., and Muniss J.N., 1972, Taxonomic value of spermatheca size for distinguishing four members of the Anopheles gambiae complex in East Africa, Bull WHO, 46: 793-799
WHO, 1975, Manual on Practical Entomology in malaria: part II, Methods and techniques, WHO offset publications, No: 13, Geneva, Switzerland
Zahar A.R.M., and Hills and Davidson G., 1970, An attempt to group fresh water species of the Anopheles gambiae complex by some morphological larval and adult characters, Parasitologia (Rome), 12: 31-46

 Author
Author  Correspondence author
Correspondence author
.png){kind=link}
{kind=link}
{kind=link}
.png){kind=link}
.png){kind=link}
.png){kind=link}